ピルビン酸デヒドロゲナーゼ複合体, PDH complex
概要: PDH complex とは
解糖 glycolysis の最終産物であるピルビン酸 pyruvate から,アセチル CoA を合成する酵素の複合体。ミトコンドリアマトリックス内に局在する(3I)。
触媒する反応は以下の通り。このサイトでは,TCA 回路の 0 段階目の反応としても説明している。

> アセチル CoA はその後 TCA 回路で酸化されるほか,脂肪酸合成などにも使われる。
> ピルビン酸は,ピルビン酸カルボキシラーゼ PC によってオキサロ酢酸としても TCA 回路に入る(補充反応)。
> この反応は好気的条件下で起こり,嫌気的条件下ではピルビン酸は LDH によって乳酸 lactate になる。
> 通常の酸素濃度にある脊椎動物の細胞では,ATP の 95% 以上が好気呼吸で産生される(4D)。
: グルコースの大部分が水と二酸化炭素になり,乳酸になるのは 4% 程度。
> ピルビン酸からアセチル CoA への反応は,3 つの酵素と 5 つの補酵素 coenzyme が必要な複雑な反応(5)。
: 3 つの酵素は PDH complex に含まれる。
: 5 つの補酵素とは thiamine pyrophosphate, lipoic acid, FAD, CoA, NAD+ である。
活性の制御
ピルビン酸からの acetyl CoA 合成は 不可逆 であり,代謝の非常に重要な分岐点である(5)。たとえば,以下のことが言える。
- 脂肪酸合成の経路は acetyl CoA から分岐し,分解された脂肪は acetyl CoA になる。これは,たとえば脂質からグルコースを合成できないことを意味する。
- いったんこの反応が起こったら,グルコースは酸化されるか脂質になるかという運命しかない。
- 細胞が低酸素状態にない,と言い切るのと同じである。
この重要な判断を下すために,PDH の活性は多くの経路によって厳密に制御されている(5)。
PDH kinase によるリン酸化制御
ミトコンドリア中のアセチル CoA/CoA 比,NADH/NAD+ 比が上がると,PDHキナーゼ(PDK)が活性化される(1I, 3I)。PDK は PDH complex をリン酸化して不活性化する(1I)。
- End product であるアセチル CoA によって PDH が不活性化される negative feedback である。
- アセチル CoA が増えることで TCA 回路が回ると,のちに酸化的リン酸化に使われる NADH が生み出される。PDH は NADH によっても不活性化される (allosteric regulation) ため,ここにも negative feedback が効いていると考えてよい。
PDH phosphatase による脱リン酸化制御
> インスリンは PDH phosphatase (PDP) を活性化する。PDP は PDH を脱リン酸化し活性化する(3I)。
> Ca2+ は PDP を活性化し,PDH を活性化させる(5)。
: 肝臓では,エピネフリンシグナルが Ca2+ シグナルを活性化させる。
代謝産物によるアロステリック制御
> Acetyl CoA 自身が PDH (E2 component) を阻害する(5)。
: 細胞で脂肪酸の分解が起こっているというシグナルである。
> NADH が PDH (E3 component) を阻害する(5)。
: TCA 回路がよく回っており,エネルギーが十分というシグナル。
> ADP とピルビン酸は,PDH kinase を不活性化することで PDH を活性化する(5)。
: これはエネルギー不足のシグナル。
脚気,水銀,亜ヒ酸
補酵素 thiamine diphosphate (ThDP) は thiamine (ビタミンB1) から合成される。ビタミン B1 不足による脚気 beriberi では,PDH の活性が低下している(5)。同様に,水銀 mercury や亜ヒ酸 arsenite も PDH を阻害する。
これらの毒物の作用は,グルコースを主要なエネルギー源とする神経系にまず現れる。水俣病のネコなど。他の組織では,グルコース以外の物質,とくに脂質をエネルギー産生に利用できるため,神経系よりも悪影響が遅く出る(5)。
MRI による in vivo での PDH flux 測定
心臓 Heart
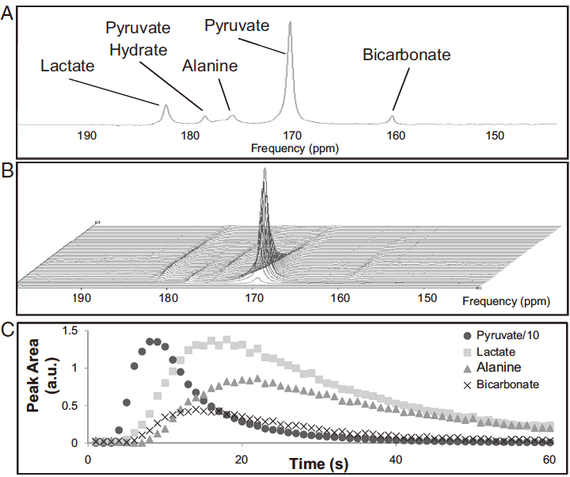
MRI (MRS) により,ピルビン酸が代謝される様子を in vivo で測定することが可能である。13C で標識したピルビン酸をラットに注射し,心臓における代謝を調べた論文がある(3R)。
13C を含む代謝産物は,下図Aのように異なる周波数のピークとして観察される。そのタイムコースをとると可視化,定量するとそれぞれ B, C のようになる。最初の大きなピルビン酸のピークが注射を示し,他の物質に代謝されていく様子をみることができる。
最終的にはTCA回路で二酸化炭素になり,炭化水素イオン bicarbonate として検出されるようになる。この論文では,絶食 fasting または streptzotocin 誘導した糖尿病のラットでは,ピルビン酸由来の HCO3- が減少することなどが明らかにされている。
つまり,飢餓または糖尿病の状態では,心臓はピルビン酸でなく他の栄養素(脂肪酸,ケトン体)を主要なエネルギー源にしていることが示唆される。糖尿病モデルでは,血糖値 blood glucose level が高いほど,ピルビン酸の代謝が低いという負の相関が示されている。
脳 Brain
> ラット脳 brain で,神経 neuron の PDH を介した代謝 flux は,1 µmol/min/g 程度である(2R)。
: 1分間に,約 1 µmol の基質から脱水素を行っているということ。
References
- Jucker et al. 1997a. 13C and 31P NME studies on the effect of increased plasma free fatty acids on intramusclular glucose metabolism in the awake rat. J Biol Chem 272, 10464-10473.
- Jiang et al. 2009a. Recurrent antecedent hypoglycemia alters neuronal oxidative metabolism in vivo. Diabetes 58, 1266-1274.
- Schroeder et al. 2008a. In vivo assessment of pyruvate dehydrogenase flux in the heart using hyperpolarized carbon-13 magnetic resonance. PNAS 105, 12051-12056.
- Lardon et al. 2005a. 1H-NMR study of the metabolome of an exceptionally anoxia tolerant vertebrate, the crucian carp (Carassius carassius). Metabolomics 9, 311-323.
- Berg et al. Biochemistry: 使っているのは 6 版ですが 7 版を紹介しています。