脳の乳酸代謝 Lactate metabolism in brain
2-21-2015 updated
- 概要
- 脳のエネルギー源としての乳酸
- CBF への影響
- ANLS
- 神経発火の調節
概要
乳酸は,血液脳関門 BBB を monocarboxylate transporter (MCT) の作用で通過することができ(1I),脳 brain には約 0.5 - 1.5 mmol/kg の濃度で存在する(2)。脳ではグルコース glucose 以外のエネルギー源として利用されるが,最近になって,受容体 HCA1 を介してシグナル分子としても作用することが分かってきた。
脳内の濃度は低く,通常 MRS では見えない(3)が,低酸素状態に曝された動物の脳では顕著なピークとして観察される。
脳での合成
> 脳内では常に合成されており,ミトコンドリアの少ないアストロサイトなどで合成量が多い(9)。
: アストロサイトも酸化的リン酸化の能力はもっているが,星状の突起は狭く,ミトコンドリアが多く存在できない。
脳への輸送
乳酸の BBB 通過は near-equilibrium transfer であり,血液中の濃度が上がると,それはそのまま脳の乳酸濃度に反映されると考えてよい(5)。逆に,脳から血液への排出も常に一定量存在する。
シグナル分子としての乳酸
乳酸は,低血糖時に脳の機能を維持する役割をもつが,これは乳酸がエネルギー源として利用されるためというよりも,乳酸が受容体 HCA1 (GPR81) を通して作用することにより,グルコースが効率的に代謝されるようになることが重要と考えられている(4R)。
なお,乳酸は低血糖時にグルコースの代わりのエネルギー源にもなっているが,それはエネルギー全体の約 10%に過ぎないことがわかっている(1I)。
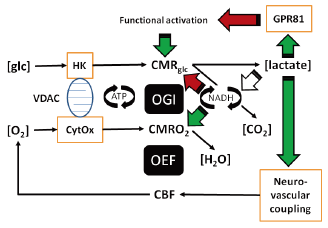
脳での乳酸の働き(5)。
> 乳酸は,エネルギー源として利用されるときに酸素を必要とする。虚血時には使われないだろう(7D)。
脳の血流 CBF への影響
Astrocyte-to-neuron lactate shuttle (ANLS)
アストロサイト astrocyte は,ニューロンとは異なり多量のグリコーゲンを細胞内にもっている(7I)。そのため,グリコーゲン分解と引き続く嫌気代謝によって,局所的にアストロサイト周辺の乳酸濃度が増大する現象が起こる。
これが,ニューロンで発火 firing のためのエネルギー源として使われることを,astrocyte-to-neuron lactate shuttle という(1I)。
> ANLSが行われているというデータはたくさんあるが,vivoでのnet transferの測定例はない(1I)。
> neuronも乳酸を放出している(1)。
> 刺激後に乳酸の濃度が局所的に上がり,5 s 後には低下することが実験的に確認されている(5)。
Lactate shuttle hypothesis
クラシックな仮説では,以下のような乳酸の機能が提唱されていた(7I)。
- アストロサイトから放出される乳酸は,トランスポーター MCT の作用でニューロンに取り込まれる。
- ニューロンでは,おそらくグルコースよりも優先してエネルギー源として使われる。
- DAB(1,4-dideoxy-1,4-imino-D-arabinitol) などでこの輸送を阻害すると,記憶に障害が現れる。
- D-乳酸も,おそらくL-乳酸と競合することで記憶を阻害する。
神経発火の調節
乳酸は,神経の発火を促進/抑制するという両方の報告がある。
> 乳酸は,マウス cortical neuron の発火を抑制する作用がある(6)。下に図を示す。
: 定量すると,約半分に低下している。濃度依存性も示されている。IC50 は約 4.2 mM。
: ピルビン酸,グルコースでは変化はわずかであるが,D-lactate でもやはり約半分になる。
: D-lactate はトランスポーター MCT との親和性が低く,細胞内に輸送される量は少ない。
: したがって,細胞外から受容体 HCA1 を介して発火を抑制していると考えられる。
: 実際に,GPCR 阻害剤 pertussis toxin で発火抑制が起こらなくなる。
: HCA1 agonist 3,5-DHBA (1 mM) でも発火抑制が起こる。
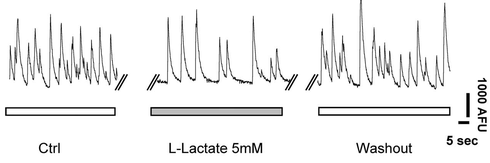
Bozzo et al. より転載(6)。初代培養したマウス cortical neuron の自発発火 spontaneous firing をカルシウムイメージングで測定。5 mM の乳酸で発火が抑制されていることがわかる。
> Astrocyte から放出される乳酸がラット locus coeruleus で発火を促進するという報告もある(8R)。
: 光刺激でアストロサイトからL-乳酸が放出されるように遺伝子改変し,発火の促進を確認。
: 外から infuse したL-乳酸でも同じ効果。D-乳酸はブロック。
: この効果は PKA-dependent であった。in vivo でも効果が合った。
: Locus coeruleus はノルエピネフリン norepinephrine 神経が集まっている部分。
: 乳酸刺激で,ノルエピネフリンの放出も促進された。
: 乳酸は 1 mM 以下なので,濃度依存的な作用が出ているのかも。HCA1 を介していない可能性も。
References
- Boumezbeur et al. 2010a. The contribution of blood lactate to brain energy metabolism in humans measured by dynamic 13C nuclear magnetic resonance spectroscopy. J Neurosci, 30, 13983-13991.
- de Graaf et al. 2003a (Review). In vivo 1H-[13C]-NMR spectroscopy of cerebral metabolism. NMR Biomed 16, 339-357.
- Govindaraju et al. 2000a. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed 13, 129-153.
- Herzog et al. 2013a. Lactate preserves neuronal metabolism and function following antecedent recurrent hypoglycemia. J Clin Invest 123, 1988-1998.
- Bergersen & Gjedde 2012a (Review). Is lactate a volume transmitter of metabolic states of the brain? Front Neuroenergetics 4, 5.
- Bozzo et al. 2013a. Lactate modulates the activity of primary cortical neurons through a receptor-mediated pathway. PLoS One 8, e71721.
- Lauritzen et al. 2013a. Lactate receptor sites link neurotransmission, neurovascular coupling, and brain energy metabolism. Cereb Cortex, published online.
- Tang et al. 2013a. Lactate-mediated glia-neuronal signaling in the mammalian brain. Nat Comn 5, 3284.
- Lauritzen et al. 2015a (Review). Monocarboxylate transporters in temporal lobe epilepsy: roles of lactate and ketogenic diet. Brain Struct Funct 220, 1-12.